Introduction
Separation of action potentials (spikes) generated by adjacent neurons is a major problem in extracellular microelectrode recording from cell-dense regions of the brain (e.g., hippocampal pyramidal cell layer). Although several types of hardware/software window discrimination have been devised, the number of neurons separable from single-channel recordings is small, averaging about two.1 Conventional procedures cannot distinguish between similar-amplitude spikes originating from different neurons equidistant from the recording microelectrode. Part of the problem has been solved by using the stereotrode method,2 which discriminates between several neurons based on the peak-amplitude ratio of spikes detected at two adjacent electrodes. Peak amplitude, however, is only one spike feature, and is unstable at low signal-to-noise (S/N) ratio or low sampling rates. Recently, a neural network method was applied to stereotrode data for multineuronal classification.3 When new multichannel spike data are to be clustered, this method requires the result of prior clustering of spike data obtained under identical conditions (the same electrode, multineuronal positions, etc.). Such a requirement imposes limitations on the applicability of the method to a variety of multichannel spike data.
We propose here a substantial improvement of the stereotrode method by using covariance, rather than just the peak-amplitude ratio, between a template and an actual spike recorded using a multisite microelectrode. Partially overlapping spikes a major obstacle to spike discrimination were separated by successively subtracting preceding spikes from raw data. Another obstacle complex spike bursts, i.e., series of spikes with different amplitudes was overcome by first detecting such bursts and then using only the first spike of each burst for spike clustering. These improvements produced better spike cluster separation. We also confirmed the correspondence between single clusters and neurons using the neurophysiological collision test. 4
Objectives
To investigate the multi-unit multisite microelectrodes embedded (implanted) in the hippocampal pyramidal cell layers of the non-primate snitch animals. If the impedance of each electrode-site is relatively low and the distance amongst electrode sites is appropriately miniatured, a spike generated by a neuron is parallelly recorded at multielectrode sites with different amplitudes.
Materials and Methods
Conventional window discriminators separate neuronal spikes by mainly referencing their peak amplitudes. However, boundaries of spike amplitudes between different neurons are difficult to determine, e.g., two neurons equidistant from the electrode site may have nearly identical amplitudes. McNaughton et al. developed the stereotrode method in which neuronal spikes are separated by referring to the joint distribution of peak-amplitude values obtained by multichannel recording.2 The method given is based on the same strategy but involves calculation of damping vectors for each observed spike. It also involves automatic decomposition of spikes that are noisy, bursty, or overlapping.
Notion
Action potentials of the target neuron are recorded extra-cellularly with a multisite microelectrode. The action potentials are assumed to originate from the center of the cell body and to exhibit consistent amplitudes at a certain distance from an active electrode site. We normalize the averaged spike waveform so that its negative peak equals
A spike generated at time T by the target neuron located at P is described as where
where
to the nth electrode site, t indicates time, and
By focusing on the attenuation effect of the spatial damping factor, (1) is transformed as
By minimization of the energy of the error function, is estimated as
Since
.....(4)
If the point of the electrode is fixed, the spatial dampening-vector
Pliable stretchy template reflects elastic
In template matching for detecting neuronal spikes,5 template waveforms should be selected, considering the variety of observed spike waveforms. Spike waveforms generated by neurons in the vicinity of a microelectrode are similar in that each contains a negative peak and a subsequent positive peak. However, spike amplitude and duration vary among different neurons.6 The differences in spike amplitude are used to determine which neuron generated a spike. Differences in spike duration make spike detection using a fixed template difficult, so we propose using an elastic template that can be fitted to different spike durations.
In preliminary spike detection based on template matching, a large spike is arbitrarily chosen as a template waveform. Spike waveforms detected in this way are then averaged. The averaged waveform is normalized so that its negative peak equals
is defined as
where Y indicates the duration ratio of
Identifying the spikes
The correlation method has been widely used for detecting neuronal spikes. In this method recorded signals having high correlation coefficients with a template are recognized as spikes.5 Detection of superimposed spikes generated by multiple neurons is a major challenge for any spike detection method. Several methods have been devised to detect such superimposed spikes. The most frequently used method is called the exhaustive search technique in which all possible combinations of predetermined templates are time-shifted to search for the best template match.7, 8 The main prob-lem of this method is the large amount of computational time required to process all possible combinations of time-shifted templates. We modify the correlation method, which is less time-consuming than the method above, such that superimposed spikes are separated by the subtraction technique described in Section II-C2.
The correlation coefficient between the observed waveform for each channel
Computing the correlation-coefficient
In order to improve the identification of the signal-spikes, the weighting-vector function can be applied for computing the correlation-coefficients. Hence, by utilizing the weight-function, the analyzed signal at the n-th electrode point
Where-in,
i.e.,
Where
Therefore, can convert the pattern-signal template as
Now we can compute the correlation-coefficient between
Therefore, we can convert the equation (11) to equation (12) by applying the equation (9), as shown in below expression
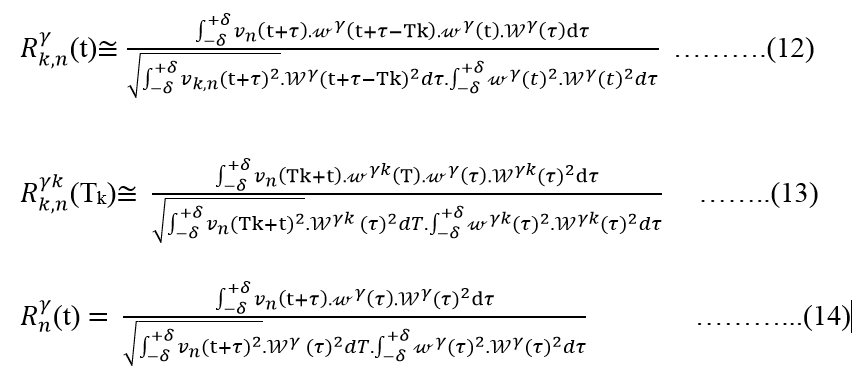
Because
2) Separation of Preceding Spikes: If the th spike and those preceding it overlap, the positive deflection of preceding spikes causes errors in estimation of damping factor
To minimize such errors, we subtract the components of preceding spikes from observed waveform
By connecting the
Computing the damping factorial
The turning points of the spike-train that are peaks acquired at various micro electrode points might have small and insignificant but then constant setbacks in some situations as multiunit data sampling synchronously (not concurrently or parallel). Say, even if the correlation function for the k-th spike observed at the i-th electrode point has a local peak value of

Hence, it is set to
Now, if
Managing burst action potential train spikes
Most subthalamic nucleus neuron jones are detected and microelectrode recording with bursting patterns can identify subthalamic neurons by their characteristic bursting pattern and their signals clearly identify the STN nucleus neurons form the surrounding structures with decreasing amplitudes, i.e., complex bursts of the spikes.9 Smaller spikes of such a complex spike burst tend to be misclassified as having been generated by different neurons. We describe here a procedure to avoid such misclassification. Since the first spike of a spike burst is considered not affected by the firing mode, the damping vectors of subsequent spikes are represented by the first spike. The subsequent spikes are then classified as belonging to the cluster of the first spike. A pair of spikes is considered to occur in a burst under the following conditions: the inter-spike-interval (ISI) is ISI b_min, ISI bax),. tTypical firing pattern with irregular firing and broad baseline noise is noticed from -1.00 level onwards…
So, they are known to fire a burst of train-spikes with reducing amplitudes, i.e., complex spike bursts
Cluster analysis
The spatial dampening vectors of the train-spikes computed/ or generated by a STN neuron are homo-centrically as a cluster in a multi-dimensional space
… cluster is characterized or embodied as a normal distribution of
Wherein ‘H’ indicating the dampening-vector as a static, ‘N’ is the height of the multi-dimensional-space equivalent to the number of electrode points, followed by the
The spatial damping vectors are clustered hierarchically by the centroid method.10 For the clustering algorithm, a Mahalanobis generalized distance is defined as the distance between the i-th and j-th clusters. The Mahalanobis generalized distance is concerned with the error rate in which damping vectors in the i-th cluster are misclassified into the j-th cluster and vice versa, i.e., the rate of the first and second type errors. 11
We have two assumptions; (1) spikes in the i-th cluster occur at the same rate as those in the j-th cluster, (2) the population variance-covariance matrix of the damping vectors included in the i-th cluster is nearly equal to that of damping vectors in the j-th cluster, i.e.,
And we can compute the error-rate such as
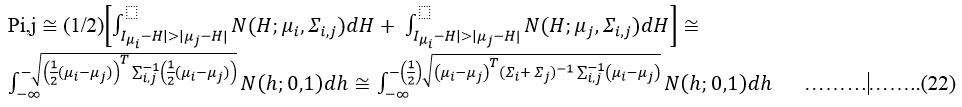
Here, H is the simplified Mahalananobis distance (statistic domain). If maximum of Pij=PIJ among all set groups (or of clusters) then i-th and j-th clusters are merged and the cluster analysis is resumed as long is PIJ
Therefore, the terminal constraint of the cluster-process is determined by the Mahalananobis statistical test. Since
Since and are unknown until the end of clustering,
Substitute values pq used. Since each damping vector initially forms a cluster, the initial value of
Results and Discussion
In this study, we understood the equivalence of a cluster-group with a subthalamic nucleus neuron. However, the validity of this notion has not yet been determined. We accomplished further investigations in which an aimed-target s t n neuron was detected by the collision-test and its damping vectors were determined by our clustering method. Since the damping vectors of the detected neuron distilled in a cluster-group which communicated well to one of the cluster-groups in the multi cluster-group allocation, demonstrating the neuron cluster correspondence. Stimulus-amplitude and pulse-widths of window discriminators of stn-neural-spikes acquired with a single site single unit microelectrode is equivalent to cluster-groups of neural-spikes including source to a minimal-distribution of damping-vectors. In distinction with single-site microelectrode recording (MER) signals of STNs, our cluster-analysis disconnected neuronal-spikes, in particular, by a joint-distribution of damping-vectors. Whether we can achieve any gain from multi-site signal-recording, nevertheless, varies on the form of multi-site multi-unit micro-electrode and on neuron-density. Neuronal-spikes computed or simulated or generated by a neuron in a cell-sparse-region, for instance, are simply separated by single-site micro-electrode recording MER signals of stn. A multi-site multi-unit microelectrode together with numerous adjoining recording-sites is necessary to distinguish neuronal-spikes computed generated/simulated by means of a number of neurons in a cell-dense-region-areas for instance the hippo-campus(campal)/pyramidal cell-layer of rascal`s. Hence, the connection among the form of multi-site multi-unit micro electrode and cell-density should be explored further than to achieve the highest advantage of use of multi-site multi-unit micro-electrode recording (MER) signals of subthalamic and hippocampal regions of the non-primate rascal animals.
Conclusion
We developed here a procedure for spontaneously categorizing several neuronal-spikes from multi-site multi-micro-electrode signals recording of stn data. The technique is vigorous sturdy and robust as compared to lower-sampling frequency and noise-distortion because a neuronal-spike is characterized as a spatial-damping-vector derived on based on the covariance-matrix amongst a pattern-matching template and the experimental signal at every single signal-recording-point of contact. The correlation among a cluster-group of damping vectors and a neuron was corroborated by the collision-test. It may be concluded that the developed procedure is useful for further accurate categorization of neuro-nal-spikes than other kinds-of-spike-sorting, which lack the techniques to distinguish corresponding-spikes or to treat spike-bursts. This developed procedure is effective in researching multi neuronal connections in locally connected neuronal-net-works.